[1]Department of Plant Pathology, and [2]USDA/ARS Plant Disease Resistance Unit, University of Wisconsin, Madison, WI 53706
*corresponding author
Email address: dkw@plantpath.wisc.edu
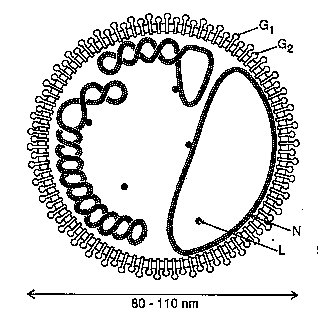
Figure 1. Representation of a tomato spotted wilt virus particle depicting the RNAs as non-covalently-closed circles associated with many copies of the N protein, G1 and G2 viral glycoproteins projecting from the membrane and a few copies of the L protein within the virion.
The ambisense 2.9 kb S RNA encodes a 52.4 kDa non-structural protein (NSs) in the viral (v) sense and the 29 kDa N protein in the viral complementary (vc) sense. The viral N protein is expressed from a viral complimentary sense 1.2 kb subgenomic mRNA and the NSs by a viral-sense sub-genomic mRNA 1.7 kb in length (De Haan et al. 1990). The 4.8 kb M RNA encodes a 33.6 kDa non-structural protein (NSm) in the v sense and a 127.4 kDa precursor to G1 and G2 in the vc sense . In an expression strategy similar to the S RNA, the open reading frames (ORFs) of the M RNA are expressed from subgenomic mRNAs of 1.1 kb (NSm) and 3.5 kb (G2G1 precursor) transcribed from opposite sense strands (Kormelink et al. 1992). The 8.9 kb L RNA is completely negative-sense and contains a single open reading frame in the vc sense encoding the 331.5 kDa L protein. L RNA expression occurs via the synthesis of a genome sized mRNA (De Haan et al. 1991; Kormelink et al. 1992).
Short, heterogeneous, non-viral sequences have been detected at the 5' ends of the mRNAs of several animal-infecting bunyaviruses demonstrating that these viruses use capped, 5' terminal sequences derived from host cell mRNAs to prime viral transcription (reviewed by Elliott 1990). These primers are generated by a process called cap snatching, in which the 5' terminal sequence of cellular mRNAs are cleaved endonucleolytically (Plotch et al. 1981). The endonuclease activity responsible for cap snatching in Germiston (Vialat and Bouloy 1992) and La Crosse (Patterson et al. 1984) bunyavirus transcription is virus-encoded.
The large size of the bunyavirus L proteins suggest that they are multifunctional, and most probably have a role in replication, transcription and cap snatching. Investigation of the putative RdRps (L proteins) of the Tospoviruses has been limited to sequence analysis of the gene encoding the TSWV L protein, which revealed the conserved polymerase motifs noted above (De Haan et al. 1991; Tordo et al. 1992), and serological demonstration of the presence of the 331.5 kDa L protein in virions (Adkins et al. 1995; Van Poelwijk et al. 1993). Non-viral sequences, 12 to 20 nucleotides in length, have recently been detected at the 5' ends of the N and NSs mRNAs of TSWV. This provides the first evidence of a plant virus using a cap snatching to prime transcription of its genome (Kormelink et al. 1992). Recent experiments in our laboratory provide the first direct evidence of an RdRp activity associated with purified virions of TSWV (Adkins et al. 1995).
Dot blot and ELISA assays. Dot blots using a TSWV-specific cDNA probe were performed as previously described (Rice et al. 1990). Enzyme-linked immunosorbent assay analysis (ELISA) was conducted using a commercially available kit (Agdia, Elkhart, IN). For both analyses, TSWV infected Datura served as a positive control.
 Click to see Fig. 2.
Click to see Fig. 2.
Figure 2. Leaves from TSWV infected and mock inoculated A. thaliana ecotype Columbia plants at three weeks post-inoculation. The chlorotic spots exhibited by the infected leaf are typical of early infection symptoms.
 Click to see Fig. 3.
Click to see Fig. 3.
Figure 3. Dot blot analysis of infected and mock inoculated A. thaliana ecotype Columbia leaves harvested from two different plants three weeks post-inoculation. Viral RNA isolated from intact TSWV virions was used as a control.
An ELISA analysis was performed in the second experiment on symptomatic leaves taken from three weeks post-inoculation. The results were as follows: TSWV inoculated = 0.687 OD490 units; mock-inoculated = 0.002 OD490 units; TSWV-infected Datura control = 0.610 OD490 units. Four weeks post-inoculation, infected plants begin to exhibit sever chlorosis and wilt. Typically the bolt will be severely chlorotic and deformed (Fig. 4). The complete wilt of infected plants prior to bolting is not unusual.
 Click to see Fig. 4.
Click to see Fig. 4.
Figure 4. Advance symptoms of TSWV on ecotype Columbia four weeks post-inoculation.
Bouloy, M. and Hannoun, C. 1976. Studies on Lumbo virus replication: I. RNA-dependent RNA polymerase associated with virions. Virology 69:258-264.
Cho, J. J., Mau, R. F. L., Mitchell, W. C., Gonsalves, D. and Yudin, L. 1987. Host list of tomato spotted wilt virus (TSWV) susceptible plants. Univ. Hawaii, Coll. Trop. Agri. Human. Resour. Res. Ext. Ser. 078,
De Haan, P., Kormelink, R., Resende, R. d. O., Van Poelwijk, F., Peters, D. and Goldbach, R. 1991. Tomato spotted wilt virus L RNA encodes a putative RNA polymerase. J. Gen. Virol. 72:2207-2216.
De Haan, P., Wagemakers, L., Peters, D. and Goldbach, R. 1990. The S RNA segment of tomato spotted wilt virus has an ambisense character. J. Gen. Virol. 71:1001-1007.
Elliott, R. M. 1990. Molecular biology of the Bunyaviridae. J. Gen. Virol. 71:501-522.
German, T. L., Ullman, D. E. and Moyer, J. W. 1992. Tospoviruses: diagnosis, molecular biology, phylogeny, and vector relationships. Annu. Rev. Phytopathol. 30:315-348.
Gonsalves, D. and Trujillo, E. E. 1986. Tomato spotted wilt virus in papaya and detection of the virus by ELISA. Plant Dis. 70:501-506.
Jin, H. and Elliott, R. M. 1991. Expression of functional Bunyamwera virus L protein by recombinant vaccinia viruses. J. Virol. 65:4182-4189.
Kormelink, R., De Haan, P., Meurs, C., Peters, D. and Goldbach, R. 1992. The nucleotide sequence of the M RNA segment of tomato spotted wilt virus, a bunyavirus with two ambisense RNA segments. J. Gen. Virol. 73:2795-2804.
Kormelink, R., Van Poelwijk, F., Peters, D. and Goldbach, R. 1992. Non-viral heterogeneous sequences at the 5' ends of tomato spotted wilt virus mRNAs. J. Gen. Virol. 73:2125-2128.
Patterson, J. L., Holloway, B. and Kolakofsky, D., . 1984. La Crosse virions contain a primer-stimulated RNA polymerase and amethylated cap-dependent endonuclease. J. Virol. 52:215-222.
Peters, D., de Avila, A. C., Kitajima, E. W., Resende, R. d. O., De Haan, P. and Goldbach, R. W. 1991. An overview of tomato spotted wilt virus: Virus-thrips-plant interaction of tomato spotted wilt virus. Proc. USDA Workshop, US Dept. of Agric. Agric. Res. Serc., ARS-87.
Plotch, S. J., Bouloy, M., Ulmanen, I. and Krug, R. M. 1981. A unique cap (m7GpppXm)-dependent influenza virion endonuclease cleaves capped RNAs to generate the primers that initiate viral RNA transcription. Cell 23:847-858.
Quadt, R., Kao, C. C., Browning, K. S., Hershberger, R. P. and Ahlquist, P. 1993. Characterization of a host protein associated with brome mosaic virus RNA-dependent RNA polymerase. Proc. Natl. Acad. Sci. USA 90:1498-1502.
Ranki, M. and Pettersson, R. F. 1975. 1975. J. Virol. 16:1420-1425.
Rice, D. J., German, T. L., Mau, R. F. L. and Fujimoto, F. M. 1990. Dot blot detection of tomato spotted wilt virus RNA in plant and thrips tissues by cDNA clones. Plant. Dis. 74:274-276.
Schmaljohn, C. S. and Dalrymple, J. M. 1983. Analysis of Hantaan virus RNA: Evidence for a new genus of Bunyaviridae. Virology 131:482-491.
Tordo, N., De Haan, P., Goldbach, R. and Poch, O. 1992. Evolution of negative-stranded RNA genomes. Sem. Virol 3:341-357.
Ullman, D. E., German, T. L., Sherwood, J. L., Westcot, D. M. and Cantone, F. A. 1993. Tospovirus replication in insect vector cells: Immunocytochemical evidence that the nonstructural protein encoded by the S RNA of tomato spotted wilt Tospovirus is present in thrips cells. Phytopathology 83:456-463.
Van Poelwijk, F., Boye, K., Oosterling, R., Peters, D. and Goldbach, R. 1993. Detection of the L protein of tomato spotted wilt virus. Virology 197:468-470.
Vialat, P. and Bouloy, M. 1992. Germiston virus transcriptase requires active 40S ribosomal subunits and utilizes capped cellular RNA. J. Virol. 66: 685-693.
Wijkamp, I., Van Lent, J., R., K., Goldbach, R. and Peters, D. 1993. Multiplication of tomato spotted wilt virus in its vector, Frankliniella occidentalis. J. Gen. Virol. 74:341-349.
 Return to Contents Page: Weeds World Vol2(i)
Return to Contents Page: Weeds World Vol2(i)